 Ако предлагате частни уроци, може да рекламирате услугите си напълно безплатно!
Ако предлагате частни уроци, може да рекламирате услугите си напълно безплатно!
ЕУКАРИОТНА КЛЕТКА
1. От прокариотна към еукариотна
клетка
1.1.От прокариотна към еукариотна клетка
Възрастта на
еукариотите все още е твърде спорна: според повечето автори между 1,5 и 2
милиарда години, според някои само около 1 милиард години. Която и да е вярната
дата, вижда се, че за появата на еукариотната клетка е било нужно много време –
може би повече, отколкото за възникването на самия живот. Следователно пътят от
прокариотна към еукариотна клетка е бил дълъг и труден.
Всички съвременни
еукариоти имат набор от общи производни (апоморфни) белези. Следователно
надцарството на еукариотите за разлика от това на прокариотите е истинска
монофилетична група, която обединява всички потомци на някакъв древен
вид-предшественик. Сравнителното разглеждане на еукариотите и различните
прокариоти сочи, че прародителят на еукариотите трябва да е бил близък до
архебактериите. Ако трябва да бъдем точни, еукариотите и архебактериите
изглеждат сестрински групи, т.е. монофилетични групи, получени чрез дивергенция
от общ предшественик.
Сравнени с прокариотите, еукариотите имат крайно
еднообразна обмяна на веществата. От това може да се заключи, първо, че още от
възникването си еукариотите са обитавали сравнително постоянна околна среда, и
второ, че еукариотната клетъчна организация за разлика от прокариотната не
насърчава големите експерименти със собствения метаболизъм. Съставът на скалните
пластове показва, че преди 2 милиарда години атмосферата вече е съдържала
молекулен кислород поради фотосинтезата на цианобактериите. Изглежда,
предшественикът на еукариотите е понасял кислорода, но по своята обмяна е бил
хетеротрофен анаероб и е извършвал ферментация. Такъв тип метаболизъм имат
например съвременните млечнокисели бактерии.
1.2.
Компартментизация
Прокариотните и еукариотните клетки донякъде
се припокриват по размери, но средно еукариотните са по-едри: най-често 10 – 100
микрометра срещу 1 – 10 микрометра за прокариотните.
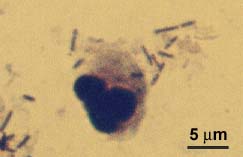
Сравнение между
прокариотна и еукариотна клетка по размери: левкоцит сред бактерии във
влагалищна намазка на пациентка. Препаратът е любезно предоставен от Димитър
Нашев от Националния център по заразни и паразитни болести.
В живия свят
уголемяването може да има неочаквани и значими последици. Разликата в размерите
често води до разлики в устройството или/и жизнените функции – явление, наречено
алометрия. Причината е, че при увеличаване на всички линейни размери
повърхността и сечението растат като квадрат, а обемът и масата – като куб. За
да се запази например отношението обем – повърхност, важно за много жизнени
процеси, трябва повърхността да се увеличи допълнително. Отличителните черти на
еукариотната клетка са й нужни за нормално функциониране при нейната големина.
Сред прокариотите най-едри клетки имат някои цианобактерии и
хемосинтезиращи бактерии. Всички те растат бавно, което показва, че са
надхвърлили оптималната големина, съответстваща на тяхната организация. И все
пак автотрофните прокариотни клетки могат да си позволят сравнително големи
размери. Техният начин на хранене не изисква усилен транспорт през мембраната.
За хетеротрофна и при това анаеробна клетка положението е различно.
Ферментацията изисква непрекъснато внасяне на голямо количество хранителни
вещества и изнасяне на недоразградените крайни продукти. При уголемяване на
такава клетка повърхността й ще расте по-бавно от обема и много скоро ще се
окаже недостатъчна.
Един начин да се увеличи повърхността е капки от
околната течност, заградени с части от клетъчната мембрана, да се включат в
цитоплазмата. Така при практически същия обем клетката ще има много по-голяма
допирна повърхност с външната среда, понеже в самата цитоплазма ще има
пространства, топологично равностойни на средата. (Топологията е клон от
математиката, който изучава деформирането на фигурите.) Всяко вътреклетъчно
пространство, оградено с мембрана, се нарича компартмент или отдел, а
обособяването на части от клетката чрез вътрешни мембрани – компартментизация
(от англ. compartmentation – подразделяне).
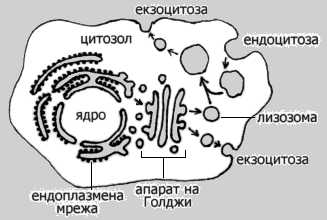
Отдели на еукариотната
клетка, топологично равностойни на външната среда. Със стрелки са означени
основните посоки на пренос на вещества. По Alberts et al. (1994) с
изменения.
Еукариотната
клетка съдържа редица мембранни органели. Всяка от тях има поне по един
компартмент, а често и повече. Мембранната преграда позволява всеки компартмент
по състав и рН оптимално да се приспособи за даден жизнен процес, без това да
засегне съседните компартменти. Така става възможно много по-голямо
разноообразие от функции, стига всяка от тях да протича на отреденото
място.
Наченки на компартментизация има и при прокариотите – да си
припомним ламелите и тилакоидите на фото- и хемосинтезиращите бактерии.
Вътреклетъчната мембранна система на прокариотите обаче не се разраства, защото
бързо би излязла извън контрол. Еукариотната клетка регулира разположението и
движенията на своите мембрани чрез съвкупност от белтъчни нишки, наречена
цитоскелет. Основните съставки на цитоскелета са микротубулите и
микрофиламентите.
Част от
мембранните органели са трайни и специализирани – ендоплазмена мрежа, апарат
(комплекс) на Голджи. Други са мехурчета със сравнително къс живот. Някои от
тях, наречени ендозоми, съдържат част от околното вещество. Те се получават чрез
вгъване на клетъчната мембрана навътре с участието на микрофиламентите –
ендоцитоза. Други мехурчета се образуват вътре в цитоплазмата и съдържат
вещества, предназначени за клетъчната мембрана или външната среда. Изнасянето им
става чрез сливане на мехурчето с клетъчната мембрана – екзоцитоза.
Противоположните и същевременно много подобни процеси екзо- и ендоцитоза общо се
наричат цитоза.
Както знаем, диагностичен белег на еукариотната клетка е
ядрото. То също се образува благодарение на съвършения контрол на клетката върху
мембраните. Обособяването на ядрото може да се опише като подреждане и частично
сливане на мембранни мехурчета около хромозомата (хромозомите). Вероятно в един
момент от еволюцията се е наложило ДНК да се загради, за да бъде защитена от
растящите механични напрежения в цитоплазмата. Вътрешността на ядрото е
топологично равностойна на цитозола.
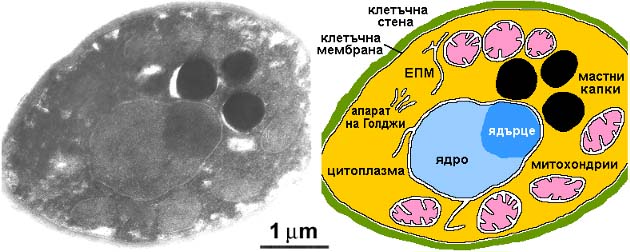
Дрождена клетка (Saccharomyces
cerevisiae) като пример за еукариотна клетъчна организация: вляво –
електронно-микроскопска снимка, любезно предоставена от Георги Милошев от
Института по молекулярна биология, вдясно – обяснителна схема. ЕПМ –
ендоплазмена мрежа.
 Наличието на ядро значително усложнява
клетъчното делене. Сравнително простото напречно делене на прокариотните клетки
е подходящо за тях, но не и за клетка с ядро. Ето защо еукариотите са
разработили митозата като начин за разпределяне на наследствения материал между
дъщерните клетки. Митозата е изключение от правилото, че цитоскелетът е изолиран
от ДНК. Хромозомите се теглят от микротубули, изградени специално за тази цел, в
някои случаи в самото ядро. Съвкупността им се нарича делително или митотично
вретено.
Наличието на ядро значително усложнява
клетъчното делене. Сравнително простото напречно делене на прокариотните клетки
е подходящо за тях, но не и за клетка с ядро. Ето защо еукариотите са
разработили митозата като начин за разпределяне на наследствения материал между
дъщерните клетки. Митозата е изключение от правилото, че цитоскелетът е изолиран
от ДНК. Хромозомите се теглят от микротубули, изградени специално за тази цел, в
някои случаи в самото ядро. Съвкупността им се нарича делително или митотично
вретено.
Усложняващата се клетъчна организация е изисквала все повече и
повече гени. Прокариотният тип опаковка на наследствения материал (една
пръстенна "хромозома") е удобен за къси ДНК-вериги, но явно неподходящ за
по-дългите. Еукариотите са преминали към линейни молекули ДНК, и то обикновено
повече от една. Вероятно митозата, въведена заедно с ядрото, впоследствие е
позволила ДНК да се разпределя успешно между дъщерните клетки дори ако е
опакована в две или повече различни хромозоми.
Обособеното ядро и
хромозомите са се оказали годни за съхранение на големи количества ДНК. Затова
еукариотната ядрена ДНК е склонна да се натрупва. Често в еволюцията на различни
групи еукариоти се удвояват отделни гени, групи гени или целият хромозомен
набор. След това размножените гени и цели хромозоми постепенно дивергират,
натрупвайки разлики в структурата и функцията си. По-свободното отношение към
собствената ДНК се проявява и в целенасочената обмяна на гени между различни
индивиди от един вид, наречена полов процес.
1.3. Фагоцитоза и
ендосимбионтни органели
След като цитоскелетът е осигурил опора
на клетъчната мембрана отвътре, външната опора (клетъчната стена) вече не е била
толкова необходима. Тя е изтъняла, станала е гъвкава и в даден момент е
изчезнала напълно. Много съвременни еукариотни клетки имат стена или обвивка над
мембраната, но това явно са по-късни образувания. Те се отличават по състав не
само от всички прокариотни клетъчни стени, а и между различните групи
еукариоти.
Меката повърхност и цитоскелетът са позволили на клетката да
се движи чрез псевдоподи като съвременните амеби. Станала е възможна типичната
фагоцитоза: обхващане и поглъщане на сравнително големи частички. Получават се
големи ендозоми, наречени фагозоми, чието съдържимо след това се смила.
Еукариотната клетка-организъм се е превърнала в хищник, способен да поглъща
по-дребните прокариотни клетки.
На този етап е протекло едно от
най-важните събития в еволюцията на еукариотната клетка: снабдяването й с
митохондрии. Еукариотите са възникнали в кислородна среда по време, когато много
от прокариотите са били строги аероби. Дори при съвременните еукариоти обаче
повечето биохимични реакции в цитозола са анаеробни. Производството на етилов
алкохол се основава на ферментация, извършвана от еукариот – Saccharomyces
cerevisiae. Нашите мускулни клетки при кислороден недостиг преминават към
обмяна, която се отличава от млечнокиселата ферментация само по името.
Молекулният кислород все пак е необходим за някои синтези в цитозола и за
окислителните реакции в пероксизомите (мембранни органели със спорен засега
произход). Но това, което прави еукариотната клетка истински аеробна, са нейните
митохондрии. Те осъществяват клетъчното дишане.
Приликата между
митохондриите и аеробните бактерии, както и между хлоропластите и
цианобактериите, се забелязва още през ХІХ в. Редица автори, например А.Ф.В.
Шимпер (1883), Р. Алтман (1890), К. Мерешковский (1905) предполагат, че
митохондриите произлизат от бактерии, а хлоропластите – от цианобактерии. Според
тази хипотеза прокариотите-предшественици някога са проникнали в еукариотната
клетка, установили са с нея взаимоизгодни отношения и с времето са загубили
способността си за самостоятелен живот. Такова отношение между два организма от
различни видове, при което единият (ендосимбионт) живее в другия (гостоприемник)
и му носи някаква полза, се нарича ендосимбиоза. Обикновено и ендосимбионтът
извлича изгода от съжителството, но в някои случаи изглежда, че просто няма как
да избяга от гостоприемника си. Ендосимбиозата е доста разпространена сред
съвременните организми – много еукариоти съдържат в клетките си прокариотни или
дребни еукариотни ендосимбионти. Не е известна ендосимбиоза с прокариот в ролята
на гостоприемника, понеже прокариотните клетки са малки и неспособни на
цитоза.
Ендосимбионтният произход на митохондриите и хлоропластите дълго
време е любопитна, но непроверима хипотеза. Постепенно с развитието на
клетъчната биология се трупат данни в нейна подкрепа. Митохондриите и пластидите
имат двойна мембрана, като външната прилича на останалите клетъчни мембрани, а
вътрешната доста се различава от тях. Това навежда на мисълта, че вътрешната
мембрана има чужд произход, а външната е мембрана на фагозома. Митохондриите и
хлоропластите имат собствена ДНК, която е пръстенна като прокариотната.
Притежават и собствени рибозоми, също от прокариотен тип. И в митохондриите, и в
аеробните бактерии дихателните ензими и АТФ-синтетазата се намират под
"мембраната" (т.е. вътрешната мембрана при митохондриите и клетъчната мембрана
при бактериите). Хлорофилът в хлоропластите се съдържа в тилакоиди, подобни на
тези в цианобактериите. Ендосимбионтната теория става общоприета след
публикуването на книгата "Произход на еукариотните клетки" от Лин Маргулис
(Margulis) през 1970.
Как трябва да са протекли събитията: Между
частичките, поглъщани от примитивната еукариотна клетка, е имало аеробни
бактерии. Някои от тях, вместо да бъдат смлени, са останали да живеят и да се
размножават в цитоплазмата. Те са се оказали полезни за клетката-гостоприемник,
понеже чрез своята аеробна обмяна са отстранявали опасния О2 и ненужните крайни
продукти на ферментацията. С течение на времето бактериите са загубили стените
си и са се оказали заградени само с две мембрани – своята и тази на някогашната
фагозома. Всяка бактериална клетка е била принудена да изнася част от
преобразуваната при дишането енергия в удобен за използване вид. Така
еукариотната клетка е станала аеробна и зависима от своя ендосимбионт. След
време част от гените на бактерията са били изтеглени в ядрото. С това тя е
престанала да бъде цялостна жива система и се е превърнала в митохондрия, както
я познаваме днес. Понеже митохондриите осигуряват енергия не само за своите
нужди, а и за цялата еукариотна клетка, вътрешната митохондриална мембрана (за
разлика от бактериалната) увеличава повърхността си чрез гънки, наречени кристи.
По-късно някои от еукариотите са погълнали прокариоти, подобни на
съвременните цианобактерии. Между двете клетки се е установила нова
ендосимбиоза. Отново гени от бактерията са преминали в ядрото. Гостоприемникът е
дал начало на еукариотните фотоавтотрофи (водораслите и зелените растения), а
ендосимбионтът – на техните хлоропласти. Така че еукариотите не само имат
еднотипна обмяна, а и са взели от прокариотите два от най-важните й процеси –
дишането и фотосинтезата.
Интересно е кога еукариотната клетка е
погълнала своите ендосимбионти и изобщо в каква последователност са се появили
апоморфните белези, отличаващи еукариотната от прокариотната клетка. Този въпрос
е труден, понеже засяга цял набор от взаимосвързани признаци, които се откриват
при всички изследвани еукариоти и липсват при всички изследвани прокариоти.
Например не е известен организъм с ядро, но без микротубули или обратното.
Логично е преходните форми да са измрели, понеже са били по-зле адаптирани от
своите еукариотни потомци, а същевременно нямат и предимствата на прокариотната
клетъчна организация. Без запазени междинни етапи обаче е трудно да се правят
изводи за историята на еукариотната клетка. По-горе нейните отличителни черти
бяха посочени в някакъв ред, но той е въведен за целите на изложението и няма
претенции да възпроизвежда действителния времеви ход на събитията.
До
неотдавна се е смятало, че митохондриите са придобити сравнително късно, след
като клетката-гостоприемник вече е развила останалите еукариотни белези.
Известни са няколко едноклетъчни еукариоти, които имат анаеробна обмяна и са
лишени от митохондрии (например паразитът Giardia lamblia). Предполагало се е,
че тези едноклетъчни са дивергирали от предшественика на останалите еукариоти,
преди той да се сдобие с митохондрии. Впоследствие обаче се разкрива, че ядрата
им съдържат гени с митохондриален произход. Следователно тези видове някога са
имали митохондрии и са ги изгубили вторично, приспособявайки се към
безкислородна среда.
Така митохондриите се оказват много по-древни,
отколкото се е предполагало, и е възможно да са се появили едновременно с ядрото
или дори преди него. Можем дори да се запитаме дали поначало ядрото не е
възникнало като място за складиране на ДНК, отмъкната оттук-оттам. Изясняването
на тези въпроси засега трябва да изчака.
Всички изследвани съвременни
митохондрии си приличат достатъчно, за да заключим, че са били придобити само
веднъж от общия предшественик на съвременните еукариоти. Прокариотният
предшественик на митохондриите най-вероятно е бил аеробен представител на
протеобактериите, към които спада и моделът E. coli.
Придобиването на
хлоропласти (или по-общо на пластиди, понеже те не винаги имат фотосинтетична
функция) изглежда значително по-късно. Освен това за разлика от митохондриите
хлоропластите явно са възникнали повече от веднъж. Отдавна е известно, че
червените водорасли по биохимията на своята фотосинтеза значително се отличават
от другите водорасли и сухоземните растения. По-подробните изследвания върху
различни фотосинтезиращи еукариоти разкриват, че пластидите им са резултат от
поне няколко независими акта на ендосимбиоза. Особено интересни са случаите,
когато ендосимбиозата е възникнала сравнително скоро. Най-добре изученият такъв
организъм е едноклетъчното двукамшичесто водорасло Cyanophora paradoxa. Всяка
негова клетка съдържа от 1 до 8 пластида, наречени цианели. Те по състав и
строеж много приличат на свободно-живеещите цианобактерии и като тях имат
муреинова клетъчна стена.
Следователно
компартментизацията е по-сложно явление, отколкото я представихме първоначално.
Докато някои компартменти са образувани чрез "просто" преразпределяне на
вътрешното пространство на клетката, други са осигурени от нейните
ендосимбионти.
Не е ясно кога и как са възникнали камшичетата, ресничките
(които са им принципно сходни) и центриолите. У еукариотите не са открити
камшичета от прокариотен тип. Дори и да са присъствали у предшественика, те биха
били безполезни за клетка с такива размери. Еукариотните камшичета са с един
порядък по-дебели от прокариотните. Всяко камшиче съдържа ос от микротубули и се
изгражда върху цилиндър от микротубули – центриола, в този случай обикновено
наричана базално телце.
Когато присъстват в клетката, центриолите
участват и в изграждането на микротубулния цитоскелет. Около тях се събира
аморфен материал, съдържащ гама-тубулин, и така се образува центрозомата. От
този материал водят началото си цитоплазмените микротубули. Затова за
организмите, които я притежават (т.е. за много едноклетъчни и за многоклетъчните
животни), центрозомата е централен микротубулен организатор.
Поначало
обаче микротубулите в цитоплазмата могат да се образуват и без центрозома. Важен
е гама-тубулинът, а не е нужно аморфният материал, който го съдържа, да е събран
около двойка центриоли. Затова не само камшичетата, а и центриолите не са
абсолютно необходими на еукариотната клетка. Много клетки и цели организми като
семенните растения са напълно лишени от тях. Не следва обаче автоматично да
допускаме, че тези структури са късна еволюционна придобивка. Напротив, дори и
да са по-млади от самите микротубули, най-вероятно те са много древни и липсата
им е вторична. Еукариотите, които изглеждат примитивни, по правило имат
центриоли и камшичета.
Цялостното сравнение между прокариотна и
еукариотна клетка показва, че придобивките на еукариотната клетка й осигуряват
възможности, но имат и цена. Цитоскелетът дава механична самостоятелност, но
хаби много енергия. Компартментизацията позволява едновременно протичане на
разнообразни процеси, но те се извършват тромаво, с висок разход на енергия и
усложнен контрол. Например секретираните белтъци не се изнасят направо, а се
синтезират в ендоплазмената мрежа и минават през комплекса на Голджи. Наличието
на ядро и хромозоми позволява много по-богата наследствена програма. От друга
страна, ядрото забавя процесите, свързани с ДНК. Транслацията не може да
започне, преди транскрипцията да приключи. Освен това еукариотните хромозоми са
пълни с некодираща ДНК, която не носи видима полза на клетката.
Еукариотите са възприели като своя адаптивна стратегия усложняването с
всичките му предимства и ограничения. Склонността към усложняване се проявява не
само в клетъчната организация, а и по-нататък в обединяването на клетките до
многоклетъчни организми. Затова в еволюцията на живия свят извън метаболизма
всичко, което ни се струва най-интересно, се е случило на еукариотите.
2. Биологичен модел: Saccharomyces serevisiae
За да се изучи еукариотната клетка, необходим е модел, който да я представя във възможно най-опростен вид. Това веднага изключва многоклетъчните еукариоти. Техните клетки винаги имат усложнения, свързани с диференцирането, а освен това трудно се култивират извън организма. Следователно моделът трябва да се търси между едноклетъчните. Той трябва да бъде просто устроен, с малко количество ДНК и брой гени, лесен за отглеждане, безвреден и, ако е възможно, в сравнително близко родство с многоклетъчните животни.
Противно на очакванията, намерен е вид, който наистина отговаря на всички тези изисквания. Той е избран не чрез активно претърсване на многообразието от едноклетъчни еукариоти, а (като E. coli) почти случайно поради връзката му с човека. Става дума за хлебните дрожди Saccharomyces cerevisiae. Те спадат към царство Fungi (Гъби), което е сестринска група на многоклетъчните животни Metazoa, т.е. най-близката група със същия ранг. Ето защо Saccharomyces ни е толкова близкородствен, колкото е възможно за организъм, отговарящ на другите посочени по-горе изисквания за минимален еукариотен модел.
Saccharomyces в превод значи "захарна гъба", а cerevisiae е латинизация на исп. cerveza – бира. S. cerevisiae е представител на групата Ascomycota (аскомицети или торбести гъби). Тя включва и други модели – плесента Neurospora, допринесла да се установи закономерността "един ген – един ензим", и тропическите дрожди Schizosaccharomyces, които също осъществяват алкохолна ферментация. Аскомицет е и плесента Penicillium, производител на първия открит антибиотик.
Човекът използва Saccharomyces за получаване на алкохол и хляб от хиляди години. Глинени плочки от Месопотамия от около 6000 г. пр. Хр. описват рецепти за бира; към 4000 г. пр. Хр. във Вавилон са приготвяли хляб, подобен на нашия. В по-ново време (от ХІХ в. насам) дрождите са в центъра на важни биологични открития. През 1837-38 трима изследователи – Ш. Каняр де ла Тур, Т. Шван и Ф.-Т. Кюцинг, независимо един от друг съобщават, че дрождите са организми, които осъществяват алкохолната ферментация. Това слага начало на дълъг спор, доколкото най-видните химици по това време смятат процеса за спонтанен и чисто химичен, катализиран от неживи "ферменти". Пастьор изследва спиртната ферментация и накланя везните към дрождената теория. През 1877 В.Ф. Кюне разграничава катализаторите, отговорни за ферментацията, от микроорганизмите, които ги произвеждат, и предлага за тези катализатори термина ензими. Думата означава "в дрожди" (от гр. ен – във, зиме – квас, мая). Това е второто важно понятие, въведено покрай Saccharomyces. Първото е самият термин "ферментация", от лат. fermentum – квас. През 1897 Е. Бюхнер за пръв път постига ферментация в безклетъчен екстракт.
Saccharomyces е първият организъм, от който е изолирана РНК. По-късно (през ХХ в.) върху него се изяснява еукариотният клетъчен цикъл. Днес дрождите се използват в генното инженерство като производители на рекомбинантни белтъци.
Дрождите и гъбите изобщо са хетеротрофи, които всмукват хранителни вещества с цялата си повърхност като хетеротрофните бактерии. Същото е и мястото, което заемат в екосистемите – сапрофити или паразити. В еволюцията си гъбите са се сдобили с твърди клетъчни стени, изградени от полизахариди (хитин или глюкани), и са загубили приспособленията си за активно движение. Клетките им съдържат централен микротубулен организатор, но той не е центрозома, а т. нар. полярно телце на вретеното. То е плоско образувание, разположено в ядрената обвивка така, че да може да образува микротубули и навън (към цитоплазмата), и към вътрешността на ядрото. При митозата делителното вретено се образува вътре в самото ядро.
Клетките на хлебните дрожди са овални с диаметър 5 – 10 микрометра. Схема и снимка на дрождена клетка беше дадена по-горе.
S. cerevisiae е много разпространен в природата. Открива се върху листата на растенията, които секретират сладък сок. Има го и върху сладките плодове, затова от тях може да се произвежда вино, без специално да се добавят дрожди. Освен това обитава червата на някои насекоми и топлокръвни животни. Хлебните дрожди са факултативни анаероби и при липса на кислород преживяват чрез алкохолна ферментация. На това се основава използването им от човека за производство на хляб и спиртни напитки.
В лабораторни условия дрождите се отглеждат лесно върху среда, съдържаща въглероден източник (обикновено глюкоза), азотен източник (обикновено амониев сулфат), витамини и минерални соли. Дрождите се нуждаят от малък брой витамини, всъщност те сами синтезират витамини от групата В толкова ефективно, че се използват като техен източник. Средата за култивиране може да бъде течна или желирана с агар. Изобщо отглеждането им в лаборатория доста напомня това на E. coli. Оптималната температура е 20 – 25оС с граници на поносимост 0 – 40оС.
Хлебните дрожди се размножават чрез делене, като клетъчното тяло се разделя по особен начин – чрез пъпкуване. Двете нови клетки не са поначало равностойни, а едната, наречена дъщерна, се образува като пъпка върху другата, наречена майчина. Отначало пъпката е съвсем малка и нараства постепенно, докато почти или напълно се изравни по размери с майчината клетка. Ядрото се дели чрез митоза, запазвайки целостта си. Едно от двете новополучени ядра преминава в дъщерната клетка, както и част от цитоплазмените органели. Между двете клетки стената расте, стеснявайки се, докато ги раздели (както при бактериите). Често Saccharomyces и другите дрожди, размножаващи се по описания начин, се наричат пъпкуващи дрожди. Има и дрожди, например род Schizosaccharomyces, при които изходната клетка не пъпкува, а се дели на две равностойни. Те се наричат "делящи се", за да се разграничат от пъпкуващите.
Истински мицел, какъвто имат повечето гъби, при дрождите не се наблюдава. При пъпкуването на Saccharomyces обаче често няколко клетки, потомци на една изходна, остават заедно известно време. В крайна сметка те се разделят и върху стената на майчината клетка остава кръгъл белег. Долната снимка показва как клетка, все още свързана с майчината, вече образува своя пъпка. Виждат се и белези от предишни пъпкувания.
Пъпкуващи клетки S. cerevisiae под сканиращ електронен микроскоп. Снимката е любезно предоставена от Marten Veenhuis от Гронингенския университет.

През по-голямата
част от жизнения си цикъл Saccharomyces е диплоиден. При определени условия се
дели чрез мейоза, давайки хаплоидни клетки. Те се разполагат в обща торбичка –
аскус, дала името на аскомицетите. По-късно хаплоидните клетки се сливат две по
две, възстановявайки диплоидното състояние. Много лабораторни щамове S.
cerevisiae обаче са с променен жизнен цикъл и основното им състояние е
хаплоидното.
Хаплоидният брой хромозоми (n) на Saccharomyces е 16. За
отбелязване е, че те не се виждат като отделни микроскопски образувания на нито
един етап от митозата. Могат да се забележат, макар и не много уплътнени, само
през мейозата. Не е ясно дали това е примитивен белег или вторично опростяване.
Други дрожди, например Schizosaccharomyces, кондензират хромозомите си по време
на митозата.
Геномът на Saccharomyces cerevisiae е разчетен изцяло. Той
съдържа почти 6000 гена, т.е. по броя на гените си Saccharomyces не се отличава
много от типичния прокариот E. coli. Изглежда, в еволюцията си хлебните дрожди
са удвоили целия си наследствен материал, и то вероятно повече от веднъж.

Коментари